


Похвальное слово катастрофам
Земные слои, образовавшиеся на месте древних морей и континентов, представляют собой единственную природную летопись, на гигантских окаменевших страницах которой записаны события далекого и близкого прошлого нашей планеты. За последние 4 млрд лет органический мир Земли прошел длинный путь от первозданных микробных сообществ до эволюционной вершины – разумного существа. Этот путь сопровождался многочисленными геологическими потрясениями, кардинально менявшими условия жизни на планете. Такие геологические «удары» по экосистемам можно рассматривать как крупномасштабные природные эксперименты, которые невозможно воспроизвести в лабораторных условиях. Во фрагментарных, но поддающихся расшифровке свидетельствах этих событий удается найти ответы на многие интригующие загадки феномена жизни, одна из которых – созидательная роль геологических катастроф в становлении современного облика биосферы
Эволюционная теория – плод союза биологии и палеонтологии
Как известно, XIX век породил эволюционное учение, ставшее впоследствии идейной основой естествознания. Представление о постепенном развитии мира вызревало у первых эволюционистов на разной «почве». У Ж. Ламарка – на биологической, как идея о возникновении биологического разнообразия. У Ч. Лайеля, который сделал акцент на изменениях Земли как субстрата жизни, – на геологической. Самый известный эволюционист – Ч. Дарвин – обобщил как биологические знания об изменчивости существующих видов, так и палеонтологические свидетельства видообразования в геологическом прошлом. Именно такой, передовой для своего времени, «междисциплинарный» подход обеспечил теории Дарвина победоносное шествие.
 Ее базовые положения, касающиеся факторов видообразования (наследственность, изменчивость, отбор) сохраняются в эволюционном учении до сих пор. Но вот по поводу конкретных механизмов этой триады сразу же возникли горячие дискуссии. Дарвина обвиняли в том, что он преувеличивал роль внешних факторов, т. е. среды, и недооценивал саморазвитие видов за счет внутренних биологических механизмов. О последних он и не мог знать, поскольку они стали известны только после открытия Г. Менделем и его последователями законов наследования и рекомбинации признаков.
Ее базовые положения, касающиеся факторов видообразования (наследственность, изменчивость, отбор) сохраняются в эволюционном учении до сих пор. Но вот по поводу конкретных механизмов этой триады сразу же возникли горячие дискуссии. Дарвина обвиняли в том, что он преувеличивал роль внешних факторов, т. е. среды, и недооценивал саморазвитие видов за счет внутренних биологических механизмов. О последних он и не мог знать, поскольку они стали известны только после открытия Г. Менделем и его последователями законов наследования и рекомбинации признаков.
В конечном итоге произошло размежевание между биологами и палеонтологами по проблеме соотношения внутренних и внешних факторов эволюции. Первые сосредоточились на изучении внутренних механизмов эволюционного процесса, вторые же – на изучении влияния внешней среды. Этот «раскол» в рядах ученых не удалось преодолеть до сих пор. Информационный «взрыв» в XX веке породил много триумфальных научных открытий, но в то же время «разбил» естествознание на множество мелких ручейков, которые нелегко собрать в единый поток. Для этого нужен новый союз биологии и геологии, совместные усилия ученых разных специальностей.
Самые выдающиеся научные открытия последарвиновского периода были связаны с проникновением в микромир живых существ. Эволюционную теорию постепенно монополизировали новые разделы биологии: цитология, генетика, молекулярная биология, биохимия. Мы узнали, каким образом за счет внутренних биологических процессов происходит микроэволюция, т. е. направленные изменения организмов на видовом и внутривидовом уровнях.
") Но микроэволюция – только первый шаг в историческом развитии живого мира. Главные результаты этих изменений проявляются в масштабе геологического времени в виде появления все новых и новых надвидовых образований (таксонов) – новых родов, семейств, типов... Без такого макроэволюционного процесса жизнь никогда бы не достигла современного многообразия. Именно резкие качественные скачки придают эволюции необратимый, поступательный характер, в то время как микроэволюция в принципе обратима.
Но микроэволюция – только первый шаг в историческом развитии живого мира. Главные результаты этих изменений проявляются в масштабе геологического времени в виде появления все новых и новых надвидовых образований (таксонов) – новых родов, семейств, типов... Без такого макроэволюционного процесса жизнь никогда бы не достигла современного многообразия. Именно резкие качественные скачки придают эволюции необратимый, поступательный характер, в то время как микроэволюция в принципе обратима.
Кстати сказать, понятия микро- и макроэволюция были введены нашим выдающимся генетиком Ю. Филипченко еще в 1927 году. Но только теперь, когда прочитаны многие страницы геолого-палеонтологической летописи, мы начали понимать, что эволюция жизни на Земле определяется эволюцией самой планеты. Именно геологические условия определяют геохимическую основу жизни, контуры жизненного пространства, направления развития жизненных форм и периодичность изменений всей биосферы в целом. Глобальная экосистема Земли как высшее звено иерархической системы органического мира задает тот «эволюционный коридор», в котором и происходит «тонкая настройка» конкретных живых сообществ.
В нашем случае Кювье увидел в геологической летописи следы глобальных катастроф и чередующихся с ними кардинальных обновлений живого мира. Дарвин же нашел доказательства непрерывного, постепенного развития видовВзаимосвязь геологических и биотических процессов особенно ярко проявляется во время планетарных катастроф. Первым обратил на это внимание знаменитый французский ученый, основатель сравнительной анатомии, палеонтолог и геолог Ж. Кювье. Но его концепция катастрофизма была отвергнута победившими эволюционистами. Одновременно к ней прочно пристал ярлык реакционной теории, якобы доказывающей многократные акты божественного творения после каждой очередной катастрофы (между прочим, сам автор теории не был причастен к этому выводу). Но теперь геология располагает доказательствами, которые позволяют реабилитировать Кювье, не отвергая при этом Дарвина.
Лицом к лицу лица не увидать...
, теплового режима (Б), и относительных масс (по сравнению с современными) углекислого газа и кислорода (В) (по Будыко и др., 1985)") В науке было немало примеров, когда на новых витках знания отвергнутые идеи возрождались, а изначально антагонистические – примирялись. Это происходило даже в такой точной науке, как физика: вспомним квантовую и волновую теории пространства, первоначально казавшиеся несовместимыми. А потом выяснилось, что обе теории вполне адекватно описывают физический мир, но разные его грани.
В науке было немало примеров, когда на новых витках знания отвергнутые идеи возрождались, а изначально антагонистические – примирялись. Это происходило даже в такой точной науке, как физика: вспомним квантовую и волновую теории пространства, первоначально казавшиеся несовместимыми. А потом выяснилось, что обе теории вполне адекватно описывают физический мир, но разные его грани.
Еще и сейчас считается, что Дарвин окончательно похоронил катастрофизм Кювье. Но на самом деле они оба правы! Просто каждый из них увидел разные «срезы» многоуровневой, иерархической системы органического мира.
Дарвин, по существу, рассматривал только микроэволюционный процесс, считая вид «кирпичиком» в самосборке надвидовых таксонов под влиянием внешней среды. Его предшественник Кювье, который не был еще обременен обилием палеонтологических данных, смог увидеть общие изменения органического мира, т. е. макроэволюцию. Примерно так же мы воспринимаем картины импрессионистов: вблизи видны лишь отдельные мазки, а сам художественный образ проступает только на расстоянии.
В наше время мы должны рассматривать эволюционный процесс не так, как прежде. То есть не как некую смену морфологических типов без учета их функциональной роли в экосистемах, а как изменение полей взаимодействий отдельных элементов больших систем. Это новое понимание эволюции выдающийся математик академик Н. Моисеев назвал «универсальным эволюционизмом». Согласно теории больших систем биосфера могла развиваться только в тесной взаимосвязи с ее земным субстратом и через определенные критические состояния, когда возникал дисбаланс между геологической средой и биотой. Таким образом, именно геологические встряски подталкивали эволюцию биосферы: вот оно, «алиби» Жака Кювье!
Гео и био – рука об руку
Главное направление эволюции биосферы определяется причинно-следственной цепочкой «сверху–вниз», а не наоборот, как это предписывалось дарвиновской идеей самосборки («снизу–вверх»). Это было обусловлено тем, что геологические процессы формируют и изменяют геохимическую базу жизни: систему взаимосвязанных биогеохимических циклов более 40 химических элементов, обеспечивающих все жизненные процессы. Они были непосредственно связаны не только с внешними оболочками Земли (литосферой, гидросферой, атмосферой), но и с глубинными зонами, откуда необходимые для жизни вещества поступают на поверхность.
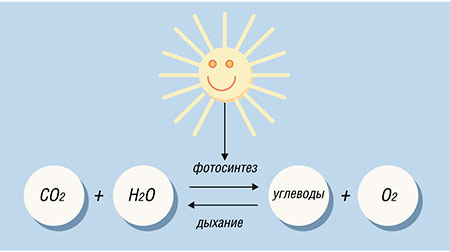 Центральную роль в функционировании глобальной экосистемы Земли играют два главных атмосферообразующих газа – кислород и углекислый газ. Непосредственное отношение к балансу этих газов имеют два процесса, осуществляемые живыми организмами – фотосинтез и дыхание. При фотосинтезе из углекислого газа и воды на свету синтезируются углеводы, т. е., по сути происходит «консервация» солнечной энергии в энергию химических связей органических соединений. При этом в качестве побочного продукта выделяется молекулярный кислород. Противоположность фотосинтезу представляет собой дыхание. В ходе этого процесса в результате окисления готовых органических веществ выделяется энергия, которая затем используется для потребностей организма.
Центральную роль в функционировании глобальной экосистемы Земли играют два главных атмосферообразующих газа – кислород и углекислый газ. Непосредственное отношение к балансу этих газов имеют два процесса, осуществляемые живыми организмами – фотосинтез и дыхание. При фотосинтезе из углекислого газа и воды на свету синтезируются углеводы, т. е., по сути происходит «консервация» солнечной энергии в энергию химических связей органических соединений. При этом в качестве побочного продукта выделяется молекулярный кислород. Противоположность фотосинтезу представляет собой дыхание. В ходе этого процесса в результате окисления готовых органических веществ выделяется энергия, которая затем используется для потребностей организма.
") При рассмотрении эволюции автотрофов (организмов, способных утилизировать углекислый газ) и гетеротрофов (использующих только готовые органические вещества) можно увидеть, что развитие первых опережало развитие вторых на всех кризисных этапах. Например, появление новых форм растительности всегда предшествовало эволюции животных. Это связано с тем, что первичным источником углекислого газа являются недра самой Земли, откуда он выносится через вулканы. Необходимые же для животных кислород и пища в виде органических веществ имеют вторичное, биогенное происхождение.
При рассмотрении эволюции автотрофов (организмов, способных утилизировать углекислый газ) и гетеротрофов (использующих только готовые органические вещества) можно увидеть, что развитие первых опережало развитие вторых на всех кризисных этапах. Например, появление новых форм растительности всегда предшествовало эволюции животных. Это связано с тем, что первичным источником углекислого газа являются недра самой Земли, откуда он выносится через вулканы. Необходимые же для животных кислород и пища в виде органических веществ имеют вторичное, биогенное происхождение.
Первичные атмосфера и гидросфера Земли были бескислородными. Понемногу, вследствие жизнедеятельности фотосинтезирующих организмов на Земле накопился свободный кислород, и вся дальнейшая эволюция жизни определялась постепенной или скачкообразной оксигенизацией воды и воздуха. Периодические вспышки вулканизма, которые особенно интенсивно проявлялись в эпохи кардинальных геодинамических перестроек (столкновения или распада континентов, образования или «закрытия» океанов и т. д.), выбрасывали в атмосферу большие порции углекислого газа и стимулировали расцвет фотоавтотрофов. Первоначально это были водные организмы, цианобактерии и водоросли. Позже, когда кислорода в атмосфере оказалось достаточно для образования защитного озонового экрана, растительная жизнь вышла на сушу. Вслед за растительностью здесь появился и животный мир.
Все население континентов по сути является эволюционным продуктом морской экосистемы. Эта гигантская биогеохимическая машина более трех миллиардов лет поставляла в атмосферу свободный кислород, которым могли дышать животные, и благодаря которому весь живой покров суши был защищен от губительного влияния ультрафиолетового излученияА теперь, чтобы не быть голословными, пролистаем геологическую летопись почти на полмиллиарда лет назад, до ордовика. На первый взгляд казалось, что этот геологический период (500–435 млн лет назад) ничем не примечателен в отличие, например, от юрского, знаменитого расцветом и последующим катастрофическим вымиранием гигантских звероящеров. Однако постепенно обнаружилось, что ордовик имеет уникальный набор совпадающих по времени биотических и геологических событий, которые в совокупности определили пути дальнейшего развития всей биосферы в целом.
Взрыв разнообразия
 Парадоксально, но первое, что было установлено, – в конце ордовика произошло одно из пяти самых крупных вымираний фауны на нашей планете. К причинам этого мы еще вернемся, сейчас важно другое. Как за деревьями не видно леса, так и этот факт долгое время заслонял другой, а именно: вымиранию организмов предшествовал самый крупномасштабный в геологической истории рост биоразнообразия – сразу втрое и в предельно короткие (по меркам эволюции) сроки!
Парадоксально, но первое, что было установлено, – в конце ордовика произошло одно из пяти самых крупных вымираний фауны на нашей планете. К причинам этого мы еще вернемся, сейчас важно другое. Как за деревьями не видно леса, так и этот факт долгое время заслонял другой, а именно: вымиранию организмов предшествовал самый крупномасштабный в геологической истории рост биоразнообразия – сразу втрое и в предельно короткие (по меркам эволюции) сроки!
На какой арене развернулись эти драматические события? Суша в то время была безжизненна, так как в атмосфере еще не было достаточного количества кислорода (примерно 10–20 % от современной концентрации) для того, чтобы образовался стабильный озоновый экран. Глубины океана также были не заселены, поскольку из-за слабого перемешивания воды там особенно остро ощущался дефицит кислорода. Подобные безжизненные зоны встречаются и в современных морях, например, в придонной части Черного моря.
Самой благоприятной для появления и развития жизни частью Земли, по крайней мере, на начальных этапах, были мелководные моря. Этому способствовало все: гидротермальные источники; доступные для солнечных лучей глубины (необходимое условие для фотосинтеза); вынос реками огромного количества минеральных веществ в растворенном и твердом виде; перемешивание воды, способствующее обогащению ее кислородом и равномерному распределению питательных веществ. Именно по этим причинам мелководные шельфовые моря до сих пор остаются самым высокопродуктивным сектором биосферы.
Вот в таких условиях и произошел в ордовике настоящий таксономический «взрыв», который поначалу оставался незамеченным. Ордовикский пик выявился неожиданно, когда были составлены временные графики изменения биоразнообразия морских организмов. Тогда американский палеонтолог Дж. Сепкоски обратил внимание на то, что близкий к современному структурный облик морской экосистемы сформировался вовсе не в кембрии (около 540–520 млн лет назад), как обычно считалось, а позже, в ордовикском периоде.
, вид сверху и снизу (ПИН РАН, Москва)")
Но что являлось причиной этого резкого скачка в биоразнообразии? Ответ был найден, когда автор этих строк провел палеоэкологический анализ ордовикской биоты.
Начало глобализации
Первое, на что нам следует обратить внимание: ордовик был одной из крупнейших эпох вулканизма за последние 500 млн лет. В это время в атмосферу Земли было выброшено огромное количество углекислого газа, необходимого для жизнедеятельности водорослей. Фитопланктон (микроскопические водоросли) появился еще в докембрии, но именно в ордовике он заселил водную толщу в массовом количестве. Расцвет фитопланктона сопровождался резким сокращением площадей циано-бактериальных «лугов», которые до этого были основным источником питания для донных животных. Произошла как бы передислокация первичного, автотрофного яруса со дна моря к его поверхности. И это не просто изменило всю пространственную структуру морской экосистемы, а вызвало ее «уплотнение», сформировало принципиально новую транспортную сеть пищевых потоков.
")
Следующим и, вероятно, наиболее важным событием было резкое увеличение концентрации кислорода. Это стало результатом быстрого роста продуктивности автотрофного яруса (водорослей), что кардинальным образом повлияло на гетеротрофное население. Почти все новые таксономические группы, возникшие в ордовике, представлены оксифильными организмами, т. е. более требовательными к концентрации кислорода по сравнению с предшественниками. Эта биологическая «улика» совпадает с геологическими данными о резкой оксигенизации поверхности Земли в этот период.
В самом деле, что нужно делать «новому» организму для выживания? Он может отправиться в эмиграцию в поисках лучшей доли, т. е. еще не занятых мест обитания. Либо осваивать новые способы добывания пищи в том же биотопе, т. е. найти свою собственную экологическую нишуЖивотное население «с благодарностью» откликнулось на кардинальное изменение ресурсной ситуации. Новообразованные группы организмов использовали появившиеся еще в кембрии основные конструктивные «наработки»: клеточную и тканевую дифференциацию, форму скелета, локомоторные устройства для передвижения и питания и т. п. На основе их рекомбинаций образовался огромный спектр новых пищевых приспособлений.
")
Одни виды способны извлекать органические вещества из ила (илоеды), другие – из частиц взвеси (фильтраторы), третьи используют растворенные питательные вещества, четвертые питаются отмершими остатками растений и животных (пастбищные и трупоядные организмы), пятые – хищники. Именно такая многозвенная трофическая сеть и начала формироваться в ордовике. С этого времени можно наблюдать постепенную глобализацию морской экосистемы: сначала на дне шельфов, затем в водной толще, и, в конце концов, в глубоководных зонах.
Многоликий ордовик
В кембрии и частично в раннем ордовике монополистами морского дна были трилобиты. Они питались илом, пропуская его через себя подобно современным дождевым червям. Кембрийский ил был особенно богат органическими веществами, так как они почти полностью попадали в осадок, поскольку в то время было мало умельцев улавливать взвешенные частицы органического вещества.
. Справа – благодаря своей форме стебельчатые иглокожие цистоидеи получили название «морских пузырей» (реконструкция)")
В ордовике появилось огромное количество фильтраторных организмов, способных питаться у дна, улавливая частицы питательных веществ прямо из воды: кораллы, мшанки, морские лилии. Наступил расцвет брахиопод, в большом количестве расплодились многочисленные виды остракод – трофических универсалов, использующих любое меню. Почти все эти организмы оказались эволюционными долгожителями и существуют до сих пор. А вот трилобиты постепенно вымерли, хотя в ордовике они еще пытались приспособиться к новым условиям. Некоторые виды трилобитов научились защищаться от хищников: одни приобрели огромные устрашающие размеры; другие стали зарываться в ил, выставляя глаза-«перископы» на длинных стебельках наружу; многие из них приобрели способность сворачиваться в своем панцире, как ежи. Однако, несмотря на все эти защитные приспособления, трилобиты оказались не в состоянии конкурировать в борьбе за пищевые ресурсы с более «продвинутыми» потребителями.
Все эти новые группы организмов составили бентос – донное или придонное население. У самого дна образовалась многоярусная структура, низший этаж которой еще занимали трилобиты, а выше – прикрепляющиеся, в основном колониальные организмы. Колониальная форма жизни, ярчайшим примером которой является современный коралловый риф, – это тоже очень эффективный способ завоевания жизненного пространства и улавливания пищи из непрерывного водного потока, порождаемого скоординированными движениями организмов.
")
Постепенно начала заселяться и вся толща воды – пелагиаль. Появились и быстро достигли расцвета первые специализированные группы пелагических организмов – граптолиты, радиолярии, возникшие еще в кембрии и первоначально предпочитавшие прикрепленный образ жизни у дна. Когда в пелагиали появилось достаточно пищи – фитопланктона – они быстро заняли эту, никем ранее не освоенную зону.
Появились многочисленные хищники: быстро плавающие наутилоидеи, первые бесчелюстные рыбы, конодонтофориды, крупные ракоскорпионы, морские звезды. В облике ордовикской морской экосистемы начали явственно проступать современные черты.
Да здравствует революция! Экологическая
, среднем (Б) и позднем (А) ордовике") Что же означало для биосферы появление столь многообразных трофических специализаций? До этого жизнь развивалась в условиях разреженного экологического пространства. При отсутствии или слабой конкуренции за ресурсы эволюционная стратегия сообществ определялась необходимостью приспособления прежде всего к нестабильной, контрастно меняющейся, абиотической («неживой») среде.
Что же означало для биосферы появление столь многообразных трофических специализаций? До этого жизнь развивалась в условиях разреженного экологического пространства. При отсутствии или слабой конкуренции за ресурсы эволюционная стратегия сообществ определялась необходимостью приспособления прежде всего к нестабильной, контрастно меняющейся, абиотической («неживой») среде.
В этой новой экологической среде, насыщенной изощренными пищевыми конкурентами, успешная эволюционная стратегия заключалась в максимальной реализации «эффекта черной королевы». «Приходится бежать со всех ног, чтобы только остаться на том же месте! Если же хочешь попасть в другое место, тогда нужно бежать по меньшей мере вдвое быстрее!» (Л. Кэрролл).
Примерно такие же процессы специализации и глобализации всей экосистемы происходят и в человеческом обществе. В пещерные времена человек почти полностью зависел от природы и в минимальной степени – от соседних племен. Если соседи становились конкурентами, люди могли уйти на поиски других угодий. Была и другая возможность – найти новую профессию: освоить земледелие либо животноводство. В современном индустриальном обществе каждый человек должен иметь собственную «экологическую нишу», свою специализацию, согласованную с другими. Согласование и компромисс – вот адекватная эволюционная стратегия в экологически насыщенных экосистемах! Такие экосистемы предложено называть когерентными. Можно считать, что именно с ордовикского периода началась когерентная эволюция, когда биотический фактор стал мощным ускорителем видообразования.
Ордовик ознаменовал новый этап в развитии жизни – этап приспособления организмов не только и не столько к физико-химическим условиям среды, которые уже начали стабилизироваться, сколько к своим партнерам по месту обитанияА что касается вымирания организмов в ордовике… Действительно, «благодаря» оледенению в конце периода произошло резкое сокращение жизненного пространства из-за осушения шельфов, что, вероятно, и послужило причиной исчезновения многих видов. Тем не менее, все основные, вновь образовавшиеся таксоны пережили оледенение, и благополучно здравствуют по сей день. И главное даже не это – важна сама необратимость перестройки всей биогеохимической системы Земли, в результате которой кардинально изменились взаимоотношения биотических компонентов друг с другом и с абиотической средой. События ордовика по сути явились настоящей экологической революцией, которая во многом определила дальнейший путь эволюции и привела к образованию миллионов разнообразных групп живых существ, составляющих непрерывный живой покров современной Земли.
В статье использованы иллюстрации из книги Е. А. Елкина и Г. М. Прашкевича «На заре жизни: БЕРЕГА АНГАРИДЫ» образцов ископаемых из коллекции ПИН РАН, г. Москва
Редакция благодарит директора ПИН РАН чл.-корр. А. Ю. Розанова за предоставленную возможность публикации материалов из коллекции ПИН РАН, г. Москва
















