


История «певчей» хромосомы
Эта статья – рассказ-«буриме», написанный шестью перьями и посвященный открытию «лишней» хромосомы в половых клетках певчих птиц. Еще не так давно такая хромосома считалась биологическим курьезом, так как была обнаружена лишь у двух бразильских видов. Сегодня, благодаря работам российских генетиков, это исключение превратилось в правило для многочисленного подотряда певчих птиц. Может быть, именно благодаря этой необычной особенности своего генома и сами эти птицы, и их певческие способности отличаются таким огромным разнообразием?
Дорожите своими исключениями!
Не прячьте их, держите их всегда на виду.
Уильям Бэтсон
Павел Бородин: Открытие исключительной хромосомы
В начале моей научной карьеры мой учитель академик Д. К. Беляев дал мне странное задание: просмотреть генетические журналы и найти там все случаи неправильного наследования признаков – исключения из законов Менделя. Д. К. тогда обдумывал идею «дремлющих» генов и искал модели для ее проверки. Я много чего нашел. Одно из исключений – неправильное наследование гена Fused у мышей – мы потом использовали как модель для пробуждения генов (Belyaev, Ruvinsky, Borodin, 1981). Другие исключения я потом описал в своей книге «Этюды о мутантах» (1983).
Наверное, сказался эффект импринтинга, но и дальше, читая научные журналы, я искал исключения. Конечно, я не мог пройти мимо истории о лишней хромосоме у зебровой амадины, которую открыли в Аргентине 21 год назад (Pigozzi, Solari, 1998).

Хромосомный набор (кариотип) клеток тела (соматических клеток) амадины хорошо изучен. У нее, как и у многих птиц и рептилий, 40 пар хромосом, из них 10 пар относительно крупных макрохромосом и 30 пар очень мелких микрохромосом. Кариотип самый обычный как для певчих птиц, так и птиц вообще.
Но лишняя хромосома зебровой амадины была совсем не обычной. Она присутствовала только в половых клетках, где была самой крупной макрохромосомой. Но в соматических клетках ее не было. Поэтому Пигоцци и Солари назвали ее хромосомой половых клеток (ХПК).
В ооцитах, предшественниках яйцеклеток, она обычно присутствовала в двух экземплярах, редко – в одном. Парные ХПК самок вели себя почти как обычные хромосомы. В начале мейоза (клеточного деления, в результате которого число хромосом уменьшается вдвое) они находили друг друга, конъюгировали по всей длине и рекомбинировали, обмениваясь друг с другом своими участками. Единственная странность была в том, что эти обмены почти всегда находились на концах хромосом, в то время как в остальных хромосомах обмены могли возникать в любой точке. У одной из самок, исследованных Пигоцци и Солари, была всего одна ХПК, которая, естественно, ни с чем не конъюгировала и не рекомбинировала.
Все самцы несли в своих сперматоцитах, предшественниках сперматозоидов, одну ХПК. Да и ту они «доносили» только до первого деления мейоза: во время или после этой стадии она вываливалась из клеток. Препараты сперматоцитов были усеяны маленькими плотными шариками – телами выброшенных ХПК. В результате в сперматозоидах эта хромосома уже отсутствовала. При этом все половые клетки, ооциты и недозревшие сперматоциты всех исследованных амадин имели хотя бы одну ХПК. Значит, они получали ее от самок. Почему же у самцов всегда одна, а у самок, как правило, две?
Пигоцци и Солари придумали сложную схему наследования ХПК, по которой она передается потомкам в одной копии через самок (Pigozzi, Solari, 2005). Согласно этой схеме, она есть в зиготе и в эмбриональных стволовых клетках, а потом все клетки, кроме предшественников половых клеток, ее утрачивают. Чтобы объяснить, откуда берется вторая ХПК у самок в ооцитах, Пигоцци и Солари предположили, что она там удваивается, а в сперматоцитах остается в одиночестве.
Научное сообщество записало эту замечательную хромосому в длинный список биологических исключений и на том успокоилось. Пять лет назад Мария Пигоцци нашла точно такую же хромосому у японской амадины (del Priore, Pigozzi, 2014). Но от этого ХПК не перестала быть исключительной – эти амадины очень близкие родственники.
Анна Торгашева: Первая ласточка
ХПК амадин так и оставалась исключением – до 25 июня 2015 г., когда я нашла ее у ласточки-береговушки. Я не искала эту хромосому. Делала свою часть проекта по анализу частоты и распределения рекомбинации в геномах птиц. Вместе с Еленой Кизиловой и Мариной Родионовой из нашего института мы выделяли из яичников птиц половые клетки на ранней стадии мейоза, готовили из них препараты и окрашивали хромосомы с использованием антител к белкам мейотических хромосом.
Хромосомы всех птиц, которых я исследовала до этого, выглядели абсолютно нормально. Эта же хромосома у самки береговушки выглядела абсолютно неправильно. Она была больше всех остальных хромосом, и она была без пары. Сначала я подумала, что это одна из половых хромосом – у некоторых птиц они образуют очень причудливые конфигурации. Но нет, у этой береговушки была нормальная пара ZW. И была эта ненормальная хромосома.
 содержит большую «лишнюю» хромосому (ХПК), которой нет в ее соматических клетках (справа). Цифрами обозначены шесть первых макрохромосом в порядке уменьшения размера, буквами Z и W – половые хромосомы. Фото Е. Башевой")
Когда я показала ее фото П. М., он возопил: «Это ХПК! Это ХПК!» – и дал статью про зебровую амадину. Наша хромосома выглядела точно так же, как ХПК амадины. Она была одной из самых больших хромосом, в одном экземпляре у самцов и обычно в двух – у самок (хотя самая первая самка имела непарную ХПК, почему я ее и заметила). Я попросила Екатерину Башеву приготовить хромосомные препараты из костного мозга береговушек. На этих препаратах большой непарной хромосомы не было. Инна Пристяжнюк сделала препараты из культуры фибробластов – не было и там. Эта хромосома была только в половых клетках. Да, это была ХПК. У ласточки-береговушки.
Любовь Малиновская: Интимная жизнь ХПК
Вскоре после открытия ХПК у береговушек я пришла в лабораторию П. М. и мне доверили детальное исследование мейоза птиц. По моим первым впечатлениям, ХПК береговушек вела себя точно так же, как ХПК амадин. У самок с двумя копиями ХПК мы наблюдали их полную конъюгацию (сближение) во время первой стадии мейоза, а точки рекомбинации (обмена) участков хромосом находились точно в тех же районах, что и у амадины, – на обоих концах ХПК.
Даже доля самок с одной копией ХПК была у них точно такой же, как у амадин (13 %), хотя береговушки были из разных природных популяций, а амадины – из зоомагазина. У всех таких самок одиночная ХПК не пыталась ни сближаться, ни рекомбинировать с хромосомами основного набора.

Поэтому когда я взялась за анализ половых клеток самцов, я не ожидала увидеть ничего нового. ХПК на препаратах распластанных ядер этих клеток часто представляет собой спутанную в клубок нить, в отличие от ровных и хорошо растянутых обычных хромосом. Порой я находила клубки вдвое большего размера, чем обычно, но списывала это на особенности приготовления препаратов. Бывает, что ХПК на разных препаратах различается по степени прокрашенности.
Сомнения стали закрадываться, когда такие клетки стали попадаться все чаще. В какой-то момент я ясно увидела два сигнала центромеры, участка, связывающего сестринские хроматиды. Не поверила, тщательно отконтрастировала фотографии и увидела две четко различимые нити, ясно указывающие на наличие не одной, а целых двух копий ХПК в одной клетке!
Сразу кричать об этом я не стала, сначала пересмотрела все предыдущие фотографии и набрала несколько десятков таких клеток. Сомнений больше не было. То, что я видела, это не особенности приготовления препаратов, не случайный «занос» копии ХПК из соседней клетки. Почти все самцы были мозаиками по числу копий ХПК. Доля сперматоцитов с двумя ХПК варьировала от 4 до 60 %.
, чижа (в центре) и береговушки (справа): все они умеют петь, и у них есть ХПК")
Откуда вообще могли взяться мозаики? Согласно модели Пигоцци и Солари и самцы, и самки наследуют одну копию ХПК от матери. У самок она удваивается во всех предшественниках половых клеток, у самцов – лишь в некоторых. Мы предполагаем, что самцы и самки наследуют одну либо две копии ХПК от матери. Самки с двумя копиями честно несут их до конца и передают своим потомкам. Самцы же, получившие от матери две копии, часто теряют одну из них в ряду обычных клеточных поколений, а другую – уже в первом делении мейоза.
Как бы то ни было, из наших данных следовало, что и у ласточек-береговушек, и у амадин ВСЕ половые клетки несут как минимум одну ХПК. У самцов она сохраняется до середины мейотического деления, у самок – до его конца. Ни одной яйцеклетки без ХПК мы не обнаружили.
Анна Торгашева: ХПК у других
 Итак, ХПК береговушки выглядела точно так же, как ХПК амадин. И вела себя точно так же. Значит ли это, что она имелась уже у общего предка береговушки и амадины, который жил 35 млн лет назад (Kumar et al., 2017)? И была унаследована в практически неизменном виде всеми его потомками? Тогда она должна быть у многих других певчих птиц, которых ни много ни мало 5 тыс. видов. Почему же ее до сих пор не видели ни у кого, кроме двух видов амадин и одного вида ласточек? Да потому, что никто ее не искал. Потому что никто никогда не анализировал их половые клетки.
Итак, ХПК береговушки выглядела точно так же, как ХПК амадин. И вела себя точно так же. Значит ли это, что она имелась уже у общего предка береговушки и амадины, который жил 35 млн лет назад (Kumar et al., 2017)? И была унаследована в практически неизменном виде всеми его потомками? Тогда она должна быть у многих других певчих птиц, которых ни много ни мало 5 тыс. видов. Почему же ее до сих пор не видели ни у кого, кроме двух видов амадин и одного вида ласточек? Да потому, что никто ее не искал. Потому что никто никогда не анализировал их половые клетки.
Мы решили это сделать и начали поиск с ближних родственников береговушки. У ее самого ближайшего – бледной береговушки – мы нашли точно такую же ХПК. Это нас не очень удивило. Эти виды разошлись относительно недавно – всего 2 млн лет назад.
Следующим из доступных для нас видов была деревенская ласточка. Хотя про доступность – это как сказать. Чтобы ее добыть, мне пришлось взбираться по хлипкой лестнице под потолок высоченного гаража, а П. М. одной рукой держал лестницу, а другой хватался за сердце. А потом вообще отошел, чтобы сделать фотографию.
Здесь нас ждало разочарование. Не под потолком, а под микроскопом. У самок деревенской ласточки все было в порядке. Никаких лишних или непарных макрохромосом мы у них не обнаружили. Это было сильным аргументом против общего происхождения ХПК у двух видов ласточек, с одной стороны, и двух видов амадин – с другой. Однако нельзя было исключить, что деревенская ласточка сама является исключением из певчих птиц и утратила свою ХПК за 18 млн лет независимой от береговушек эволюции. Нужно было продолжить поиск в соседних ветвях родословного древа.
Большинство птиц для анализа мы получали от Елены Шнайдер из Сибэкоцентра. Это были погибшие или смертельно раненые птицы, которые попадали в их Центр реабилитации птиц. Весной 2016 г. Лена принесла нам погибшего самца снегиря. И в его половых клетках мы обнаружили ХПК. Она опять была самая большая и не имела пары. То есть точно такая же, как у амадин и береговушек. Но снегирь принадлежит совсем к другой ветви птичьего древа и практически одинаково эволюционно далек и от ласточек, и от амадин.
Стало ясно, что мы поторопились хоронить гипотезу общего происхождения. И мы начали ее систематическую проверку. Хотя какая уж там систематичность. Мы просто анализировали хромосомы всех птиц, которых нам приносили. И у большинства певчих птиц мы находили крупную ХПК, которой не было в их соматических клетках.
Однако у некоторых видов никакой дополнительной макрохромосомы в половых клетках не было. И распределены по филогенетическому древу они были без всякой логики. Мы не нашли ХПК у деревенской ласточки, а также грача, щегла, канарейки, овсянки и мухоловки. Это могло означать, что ХПК возникла единожды у общего предка всех певчих птиц, но некоторые виды ее потом утратили в процессе независимой эволюции. И ХПК не является такой уж необходимой и важной.
, но в клетках костного мозга этих птиц она отсутствует (б). В отличие от самок, у самцов не имеет пары и окружена синим облаком антицентромерных антител. Цифрами обозначены макрохромосомы в порядке уменьшения размера, буквами Z и W – половые хромосомы. Фото Л. Малиновской и И. Пристяжнюк")
И тут, в очередной раз ломая голову над тем, что может быть общего у видов без ХПК, я вдруг поняла, что мы анализировали хромосомы только самок этих видов. А у самок, напомню, ХПК ведет себя в мейозе как самая обычная пара хромосом: конъюгирует, рекомбинирует… Что означает, что будь она любого другого размера, кроме самого большого, отличить ее от других спаренных микрохромосом будет невозможно. Но у самцов она будет заметной благодаря своей непарности, отсутствию сигналов рекомбинации и характерному диффузному облаку хроматина, с которым почему-то (мы пока не знаем почему) связываются антитела против центромер.
Тогда мы срочно стали искать самца деревенской ласточки. Поймать птицу удалось в одном заброшенном кирпичном бараке. И?! Да, в сперматоцитах деревенской ласточки мы увидели непарную, очень маленькую хромосому. Нерекомбинирующую, окруженную облаком антител. ХПК.
Такая же маленькая ХПК обнаружилась и у мухоловки, щегла, канарейки, овсянки, грача. Мы нашли ХПК у всех исследованных нами 14 видов певчих птиц. Плюс ХПК двух видов амадин, изученных до нас.
Итог: ХПК есть у 16 видов певчих птиц из 9 разных семейств. Вероятно, она есть у всех остальных 5 тыс. видов певчих птиц. Певчие птицы – это подотряд отряда воробьинообразных (Passeriformes). В этот отряд входит также подотряд кричащих воробьинообразных, или тиранов (Tyranni), насчитывающий около 1 тыс. видов, обитающих в основном в Южной Америке. Пути эволюции певчих и кричащих разошлись около 50 млн лет назад. Пока нам не удалось добыть ни одной «кричащей» птицы, поэтому мы не знаем, есть ли у них ХПК. А что известно о других птицах? Может, и у них есть ХПК, просто ее там до сих пор не нашли?
 есть ХПК. У одних видов она большая (закрашенный кружок), у других – маленькая (незакрашенный кружок), но есть у всех. Ни у одной исследованной птицы за пределами отряда воробьинообразных (кружок отсутствует) ХПК нет")
Мы внимательно рассмотрели все опубликованные фотографии мейотических хромосом у самцов птиц из разных отрядов. ХПК определенно нет у кур, перепелов и голубей. Эти три вида подробно проанализировала еще Мария Пигоцци, и она бы увидела ХПК, если бы они там были. Мы изучили сперматоциты гуся, крачки, стрижа, сокола-чеглока и, наконец, волнистого попугая (попугаев считают ближайшими родственниками воробьинообразных). Ни у кого из них нет ничего похожего на ХПК.
Итак, ХПК не оказалось ни у одного из десяти исследованных видов, принадлежащих к пяти другим отрядам. Следовательно, она возникла один раз у общего предка всех певчих птиц, т. е. около 35 млн лет назад, либо около 50 млн лет назад у общего предка всех воробьиноообразных (Kumar et al., 2017). Что с ней происходило в течение этого времени? Мы знаем, что она меняла форму и размеры. Но как менялось ее содержание?

Нам нужен был молекулярный зонд для сравнения генетического содержания ХПК разных видов. За ним мы пошли через коридор к Николаю Борисовичу Рубцову, специалисту в микрохирургии хромосом.
Николай Рубцов: Зонд на ХПК
В своих исследованиях организации и эволюции хромосом меня всегда тянуло к исключениям. У человека – к сверхчисленным малым маркерным хромосомам, у других видов эукариот – к дополнительным В-хромосомам, вообще к различным проявлениям хромосомной нестабильности. Предложение П. М. вместе разобраться в том, что представляют собой ХПК, вызвало живой интерес. Я сразу подключился к изучению найденных ХПК с помощью отработанных в нашей лаборатории технологий микродиссекции хромосом и создания ДНК-библиотек из индивидуальных хромосом и хромосомных районов.
Сбор материала ХПК оказался не очень сложным делом. Обычно мне приходится проникать внутрь ядра, выбирать нужную хромосому или ее отдельный район, да так, чтобы при этом не зацепить соседние хромосомы. В случае ХПК я искал клетки, в которых она уже была выброшена из ядра и лежала рядом в виде шарика. С помощью электронного микроманипулятора иглой сдвигал ХПК на свободное место, подцеплял ее и под контролем микроскопа переносил в каплю специального раствора в носике пипетки, а затем в пробирку.
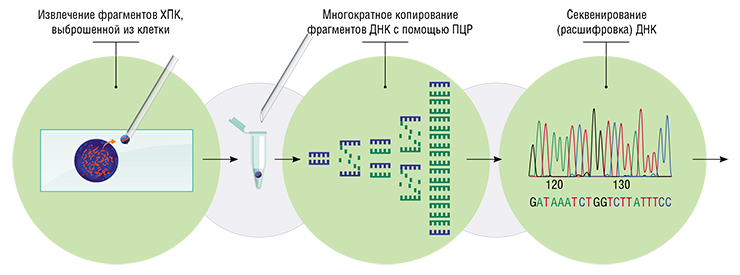
Из этого материала мои коллеги Кира Задесенец и Татьяна Карамышева выделяли ДНК и амплифицировали (многократно копировали) ее с помощью полимеразной цепной реакции (ПЦР). Полученный материал мы использовали для секвенирования и анализа генетического содержания ХПК. Из этого же материала сделали ДНК-зонд, который представлял собой набор небольших фрагментов ДНК этой хромосомы размером около 400 пар нуклеотидов, представленных миллионами копий. Эти фрагменты были помечены флуорохромами (красителями с люминисцентными свойствами).
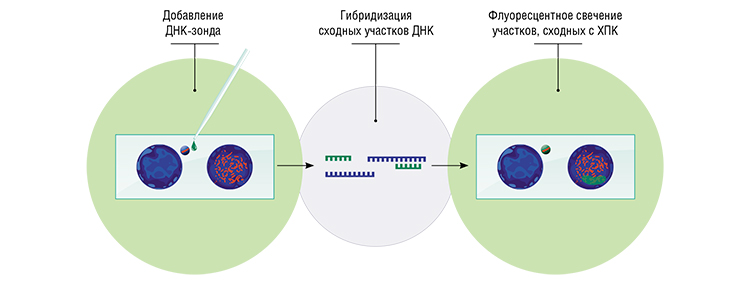
Таким образом у нас в руках оказались зонды к ХПК четырех видов птиц (зебровой и японской амадин, бледной береговушки и чижа), которые можно было наносить на препараты хромосом разных видов. После денатурации и ренатурации меченые фрагменты ХПК должны были связываться (гибридизоваться) с гомологичными («родственными») им районами хромосом образца. Эти районы можно увидеть, используя люминесцентный микроскоп.
Зонды предназначались как для опознания ХПК в любом состоянии, в любой клетке, на любой стадии клеточного цикла, так и для анализа ее генетического состава. С их помощью можно было сравнивать ХПК разных видов, искать гомологии между ХПК и участками основного генома.
. И гораздо хуже – зонд на ХПК японской амадины (б). Чем больше эволюционная дистанция между видами, тем ниже сходство между их ХПК и, соответственно, процент гибридизации. Зонд гибридизуется не только с ХПК, но и с отдельными районами хромосом основного набора, где имеются исходные копии последовательностей, находящихся в ХПК (например, на 3-й хромосоме). Фото К. Задесенец и Т. Карамышевой")
Как и ожидалось, ДНК выброшенной ХПК гибридизовалась с ХПК, находящейся внутри половых клеток. Кроме того, мы наблюдали явные сигналы гибридизации с некоторыми районами хромосом основного набора. Это означало, что в ХПК имеются копии этой ДНК. Интересно, что ХПК разных видов содержали копии из разных мест генома.
Да и в целом ХПК сильно отличались друг от друга по генетическому составу. Так, генетический материал ХПК зебровой амадины гибридизовался лишь с половиной ХПК японской амадины и почти совсем не связывался с ХПК ласточек. Это означало, что между ними очень мало сходства. Но, с другой стороны, ДНК-зонд большой ХПК бледной береговушки метил маленькую ХПК деревенской ласточки, что доказывает их общее происхождение. Этому мы были особенно рады, потому что получили наконец молекулярное доказательство, что маленькие ХПК – это таки ХПК.
Светлана Галкина: «Ламповые щетки» из ХПК
Летом 2015 г. на кафедре генетики Санкт-Петербургского университета зазвонил телефон. Это был П. М.: «Мы нашли у ласточек-береговушек ХПК. Только никому про это не говорите! Она такая же, как у амадины. И Николай Борисович сделал нам зонды на ХПК и ласточки, и амадины. Было бы замечательно посмотреть, как она выглядит в растущих ооцитах и что там она делает».
Про дополнительную хромосому в половых клетках зебровой амадины я узнала довольно давно, поскольку область моего научного интереса – организация хромосом в ядрах растущих ооцитов птиц. У птиц ооцит в процессе роста накапливает желток и дорастает до гигантских размеров – становится яйцом. В начале его созревания на хромосомах синтезируется огромное количество РНК – молекул нуклеиновых кислот, служащих матрицами для производства белков либо выполняющих регуляторную функцию.
Хромосомы при этом становятся похожими на ершики для мытья посуды – так в XIX в. выглядели щетки для мытья керосиновых ламп, поэтому с момента первого описания их называют хромосомами типа ламповых щеток. На этой стадии они достигают гигантских (по меркам клеточного мира) размеров – до одной-двух десятых миллиметра. Ими можно манипулировать при небольшом увеличении микроскопа и вручную. Методика приготовления препаратов хромосом-ламповых щеток птиц технически не очень сложна, но требует опытных рук и наметанного глаза, чтобы среди желтка различить прозрачное ядро, окруженное лишь слегка опалесцирующей ядерной оболочкой.
 (а). ХПК в ядре ооцита на стадии ламповых щеток гибридизуется с ДНК зонда на ХПК, давая зеленое свечение (б)")
Первые препараты «ламповых щеток» из ооцитов амадин мы приготовили еще в 2010 г. На них мы увидели самую большую хромосому и предположили, что это и есть ХПК. Конечно, мы ожидали, что ХПК на этой стадии должна как-то выделяться своим внешним видом – ведь она же особенная! Но оказалось, что эта пара самых больших хромосом на стадии ламповых щеток выглядит совершенно так, как обычные хромосомы на этой стадии развития ооцита. Как доказать, что это именно ХПК? Препараты сфотографировали, а файлы с фотографиями бездоказательной хромосомы сложили в архив – оказалось, на шесть лет. Нам нужен был зонд на эту хромосому, но тогда у нас его не было.
Когда Н. Б. прислал нам свой зонд, мы с Алсу Сайфитдиновой и Лерой Володькиной сделали новые препараты и провели его гибридизацию на препарате амадины. Мы получили прямое доказательство того, что та подозрительная хромосома действительно является ХПК на стадии ламповых щеток и ведет себя как любая другая хромосома на этой стадии – активно считывает информацию с ДНК на РНК. Но вот какие конкретно генетические последовательности?
Анна Торгашева: Из чего состоит ХПК
Перед нами встал главный вопрос: из чего состоит ХПК? Что за гены в ней находятся? Что там еще, кроме генов? Как геном ХПК соотносится с основным геномом? Насколько сходны и в чем различны ХПК разных видов? Чтобы ответить на эти вопросы, мы секвенировали ДНК, которую Николай Борисович добыл из выброшенных ХПК зебровой и японской амадин, чижа и бледной береговушки.
К сожалению, особенности этого метода таковы, что в анализ попадают не все участки ДНК хромосомы. Поэтому получить ее полную последовательность невозможно. Тем более, как мы уже знали из результатов перекрестной гибридизации, ХПК содержит много повторенных участков. А это дополнительно осложняет задачу.
Поэтому мы стали сравнивать последовательности, полученные в результате секвенирования, с известными последовательностями генома зебровой амадины. Таким способом мы, конечно, не могли узнать, есть ли в ХПК уникальные гены. Но зато могли увидеть то общее, что есть у нее с ДНК хромосом основного набора.
Выяснилось, что ХПК каждого вида содержит копии небольших (до 2,5 тыс. пар оснований) участков основного генома, повторенные от десятков до нескольких тысяч раз. И у каждого вида разные. Очень похоже на то, что мы видели при гибридизации. Вероятно, в разных линиях птиц в ХПК независимо размножились разные последовательности, когда-то туда скопированные.
Что же было в этих районах? К сожалению, ничего особенного. В основном некодирующие участки генов (интроны) и межгенные области. Впрочем, кодирующие участки тоже попадались. Но значительно реже и не целиком.
Денис Ларкин: В свете эволюции птичьего генома
Говорить, что птичий геном «уныло консервативный», это все равно, что назвать Солнце скучным. Пусть на свете есть миллиарды других звезд, и многие из них светят даже лучше, но только толку нам от этого чуть. Так и с геномом птиц. Действительно, по сравнению с «сумасшедшими» геномами млекопитающих у птиц он консервативен и степенен. С другой стороны, если ваш геном настолько эффективен, что позволяет вам летать (наяву, а не во сне), то зачем его сильно менять? Так ведь можно и упасть.
Чтобы научиться летать, птицам пришлось пожертвовать большей частью мобильных генетических элементов, а именно они являются драйверами эволюции у животных (Kazazian, 2004). У млекопитающих эти мобильные элементы чего только ни делают: и новые гены формируют, и регуляцию старых меняют, и хромосомы перестраивают. И все это для того, чтобы млекопитающие могли выживать и адаптироваться к постоянно меняющимся условиям среды. Кто не смог – извини.
Со стороны может показаться, что в геноме птиц по сравнению с млекопитающими ничего не происходит. Но это не так. Эволюционный процесс не останавливается, потому что остановка – это немедленная (по меркам эволюции примерно за 1 млн лет) гибель. Уместно вспомнить слова, сказанные Алисе Черной Королевой: «Здесь приходится бежать со всех ног, чтобы только остаться на том же месте! Если же хочешь попасть в другое место, тогда нужно бежать по меньшей мере вдвое быстрее». Так и с эволюцией. Приходится постоянно меняться только для того, чтобы казаться неизменным в изменчивом мире. А чтобы измениться, надо делать это еще быстрее.
Вот здесь и возникает некий птичий парадокс: чтобы летать, лучше не меняться, но чтобы не исчезнуть с лица Земли, меняться надо. Надо сказать, что птицы хорошо усвоили урок своих старших братьев – вымерших динозавров. Чтобы выжить в неблагоприятных условиях, нужно быть маленькими и по возможности не преодолевать (адаптироваться), а убегать (улетать) от серьезных трудностей. Но тогда возникает риск проиграть уже не окружающей среде, а другим животным: по суровым правилам эволюции два вида не могут обитать в одной и той же экологической нише. Кому-то обязательно придется потесниться.
Если посмотреть на все разнообразие птиц, то окажется, что из примерно 10 тыс. видов половина – это птицы певчие. Поэтому очень бы хотелось связать ХПК с таким изобилием.
, и она увеличивалась в размерах. В некоторых линиях увеличения не происходило, или оно поворачивало вспять за счет делеций (потерь)")
Мы с коллегами давно установили, что у млекопитающих в местах хромосом, в которых их сумасшедшие геномы рвутся, чтобы образовать новые структуры (и, соответственно, новые виды), находятся гены, явно связанные с наиболее выдающимися особенностями этих видов. Например, у жвачных это гены-дефенсины, защищающие их от токсинов, которые производят мириады микробов, обитающих в рубце. У свиней – гены, отвечающие за вкусовое восприятие, благодаря чему этих животных можно кормить любой малопривлекательной едой, а они будут только довольно хрюкать и жиреть.
Теперь мы знаем, что в районах таких хромосомных перестроек у млекопитающих накапливаются мобильные генетические элементы, которые меняют экспрессию нужных генов (конечно же, абсолютно случайно, но естественный отбор не дремлет). У видов, у которых подобных перестроек нет, этого не происходит.
У птиц мобильных элементов мало, и менять им что-то сложнее. Поэтому когда П. М. сказал мне, что у него есть «расшифровки» фрагментов этой необычной хромосомы, моя первая мысль была проверить, не является ли ХПК кладбищем хромосомных «ошметков», которые образуются при перестройке хромосом. В этом случае они должны были быть похожи на ДНК из районов перестроек в предковых геномах или тех же, но неперестроенных районов геномов других птиц. Оказалось, что сходства нет. По крайней мере, у зебровой амадины, а других геномов у нас не было.
Правда, выяснилась другая интересная деталь: даже у близких видов последовательности ХПК сильно различаются. Кроме того, в ХПК много тех мобильных элементов, которые были активны у предка певчих птиц, а мобильных элементов, которые были активны у общего предка всех птиц, – совсем мало. Это позволило нам датировать происхождение этой хромосомы еще одним способом. Ее, очевидно, не было у предка всех птиц, а немногочисленные древние мобильные элементы, по-видимому, перенеслись в ХПК из соматического генома вместе с другими генными последовательностями. А вот у общего предка певчих птиц она уже была.
Но остается открытым вопрос: как повлияла же ХПК на видообразование у певчих птиц? Ее большое разнообразие у разных видов птиц говорит о том, что певчие нашли отдушину, спасающую их от «унылости» основного генома, при этом его не трогая. Хотя недавно мы обнаружили, что примерно в то же время, когда появилась ХПК (около 40—35 млн лет назад), у певчих птиц вдруг начали значительно перестраиваться богатые генами маленькие хромосомы. Может, это сказалось влияние ХПК с ее «дублированными» кусками генома? Это нам еще только предстоит открыть. Или нет.
Павел Бородин: Как мы писали статью про ХПК
Первую ласточку мы «открыли» в июне 2015 г. К маю 2018 г. у нас уже был готов весь материал – пора было оформлять его в хорошую статью. На самом деле писать статью мы начали еще осенью 2016 г., когда у нас было всего две ласточки – береговушка и бледная береговушка, и первые результаты перекрестной гибридизации ДНК между ними и амадинами. Тогда мы твердо верили в недавнее и независимое возникновение ХПК в этих двух филогенетических линиях. А затем по мере поступления новых данных мы эту статью неспешно переписывали.
И все эти годы, просыпаясь по утрам, я первым делом лез в Гугл и набирал GRC songbirds. Меня не оставляло предчувствие, что нас опередят и кто-то обязательно опубликует статью под называнием «А вы знаете, ХПК есть не только у зебровой амадины…».
. Фото снегиря для обложки нам подарила Елена Шнайдер. Публикуется с согласия издателя")
3 мая 2018 г., когда у нас была написана версия № 33, в журнале Current Biology появилась статья американских исследователей под названием Discovery of the First Germline-Restricted Gene by Subtractive Transcriptomic Analysis in the Zebra Finch, Taeniopygia guttata («Открытие первого гена ХПК зебровой амадины путем вычитательного анализа транскриптома») (Biederman et al., 2018). Авторы этой статьи обнаружили на ХПК зебровой амадины ген под названием aSNAP, а в основном геноме – исходную копию этого гена, которая кодирует белок, участвующий в процессе слияния плазматических мембран.
Зачем этот ген нужен ХПК – неизвестно, но его отличия от исходной копии оказались весьма значительны. Значит, он попал в ХПК очень давно. Последняя фраза этой статьи звучала так: «Из наших данных следует, что ХПК относительно стара и может присутствовать в большем числе филогенетических линий птиц, чем первоначально ожидалось». Как написал Дарвин в схожей ситуации: «Статья Уоллеса содержала ту же теорию, что и моя».
Поэтому, не дожидаясь советов Гукера с Лайелем, мы все бросили и за два месяца дописали наконец нашу статью. Как честные люди, мы отправили ее в тот же журнал, который опубликовал наше «письмо Уоллеса»: в Current Biology. Но они даже не послали ее на рецензию, а вернули через два дня, порекомендовав отправить в более специализированный журнал. Видимо, решив, что для их общебиологического журнала одной статьи про ХПК хватит на годы вперед.
Конечно, мы не стали посылать свое открытие в специализированный журнал. Еще чего. Она явно заслуживала общечеловеческого внимания. Но наш главный секрет был раскрыт уже в названии статьи – ХПК широко распространена среди певчих птиц. Поэтому мы немедленно загрузили статью на сайт препринтов bioRχiv.org. После этого мы составили список журналов, подходящих для публикации нашего открытия, и приступили к рассылке писем редакторам.
Пока мы коллекционировали отказы от разных журналов, на том же сайте bioRχiv.org появился препринт шведско-голландско-американо-германо-испанской группы под названием Programmed DNA elimination of germline development genes in songbirds («Запрограммированная элиминация ДНК генов, участвующих в развитии клеток зародышевой линии у певчих птиц») (Kinsella et al., 2018).
Авторы этой статьи обнаружили, что:
• ХПК зебровой амадины по минимальным оценкам несет больше сотни генов, треть из которых в основном работает в яичниках, и только шесть – в семенниках. Продукты этих генов участвуют в процессах развития репродуктивной системы, особенно женской. А также в процессах клеточного развития, в том числе в дифференцировке нейронов.
• Исходные копии всех найденных генов ХПК присутствовали в основном геноме зебровой амадины и других птиц.
• Среди некоторых генов ХПК зебровой амадины есть два, которые, вероятно, были скопированы туда очень давно, еще до возникновения современного разнообразия певчих птиц, и с тех пор сильно изменились.
Из последнего следовало, что «возраст ХПК составляет десятки миллионов лет и она, вероятно, присутствует в геномах певчих птиц, что хорошо согласуется с недавним цитогенетическим препринтом». Это они ссылались на наш препринт, что с их стороны было честно и благородно.
Что меня больше всего восхищает в этой истории, так это то, что три независимые исследовательские группы практически одновременно с разных точек зрения и на разном материале пришли к одному и тому же заключению: ХПК широко распространена среди певчих птиц. Именно под таким названием в июне 2019 г., т. е. ровно через 4 года после «первой ласточки», нашу статью напечатал один из ведущих международных мультидисциплинарных журналов – Proceedings of the National Academy of Sciences (он же PNAS) (Torgasheva et al., 2019). И вынес ее анонс на обложку.
Когда вышла наша статья, к нам пришла слава. Про нас написали New Scientist, GenomeWeb, N+1, «Чердак», ТАСС, «Российская газета», «Газета.ру» и даже само «Пикабу». Но, как справедливо заметил О. И. Бендер, слава – это яркая заплата на ветхом рубище певца. Из прессы мы узнали про себя много нового. Оказалось, мы открыли, что певчие птицы поют исключительно из-за нашей исключительной хромосомы, а без нее они бы не запели. А еще – что самки птиц от того и самки, что у них в яйцах есть лишняя хромосома.
Павел Бородин и Анна Торгашева: Вопросы на завтра
И, наконец, короткий список вопросов к ХПК.
Во-первых, когда, почему и как она выбрасывается из соматических клеток?
 • Когда: насколько рано в ходе онтогенеза это происходит? Науке известно несколько случаев выбрасывания целых хромосом или их отдельных кусков из соматических клеток. Это явление под названиями диминуция хроматина и программируемая элиминация ДНК известно уже более ста лет. Оно описано у многих видов: аскарид, циклопов, листовых комариков, миног, миксин. В тех случаях, когда этот процесс исследовали, оказалось, что он происходит очень рано, иногда при самых первых делениях зиготы. Про птиц мы пока не знаем ничего.
• Когда: насколько рано в ходе онтогенеза это происходит? Науке известно несколько случаев выбрасывания целых хромосом или их отдельных кусков из соматических клеток. Это явление под названиями диминуция хроматина и программируемая элиминация ДНК известно уже более ста лет. Оно описано у многих видов: аскарид, циклопов, листовых комариков, миног, миксин. В тех случаях, когда этот процесс исследовали, оказалось, что он происходит очень рано, иногда при самых первых делениях зиготы. Про птиц мы пока не знаем ничего.
• Почему (каков в этом адаптивный смысл): чтобы уменьшить размер генома соматических клеток и расходы на его поддержание? Чтобы удалить из соматических клеток гены, необходимые для раннего развития и бесполезные или даже вредные во взрослом организме? Может ХПК – это что-то вроде загрузочного диска?
• Как: ХПК пассивно отстает от других хромосом в ходе клеточных делений? Потому, что у нее хилая центромера и к ней плохо лепятся нити веретена деления? Или клетки активно выпихивают ее из ядра?
Во-вторых, когда она возникла? Мы знаем, что ХПК есть у всех певчих птиц и что ее нет у птиц за пределами отряда воробьинообразные. Но мы не знаем, есть ли она у кричащих воробьинообразных. Эти птицы населяют экзотические страны – Австралию, Южную Америку, Юго-Восточную Азию. В Сибирь они не залетают, в наших зоомагазинах их тоже нет. Если у них нет ХПК, это значит, что она возникла у общего предка всех певчих птиц около 35 млн лет назад. Если есть, это указывает на общего предка всех воробьинообразных и старит ХПК на 20 млн лет.
И, наконец, в‑третьих: не эта ли исключительная во всех смыслах хромосома обеспечила феерическое разнообразие видов воробьинообразных птиц и быструю эволюцию их форм и песен, прекрасных и удивительных? И если да, то как?
Вместо эпилога
Уильям Бэтсон: «Дорожите своими исключениями! Исключения похожи на грубую кирпичную кладку растущего здания, которая говорит о том, что предстоит еще многое сделать, и показывает, где будет следующая постройка»
Литература
Бородин П. M. Этюды о мутантах. М.: Знание, 1983. 112 с.
Bateson W. The Method and Scope of Genetics. 1908. 45 p.
Belyaev D., Ruvinsky A. & Borodin P. Inheritance of alternative states of the fused gene in mice // Journal of Heredity. 1981. V. 72. P. 107–112.
Biederman M. K., Nelson M. M., Asalone K. C. et al. Discovery of the First Germline-Restricted Gene by Subtractive Transcriptomic Analysis in the Zebra Finch, Taeniopygia guttata // Curr Biol. 2018. V. 28. N. 10. P. 1620–1627.
del Priore L. & Pigozzi M. I. Histone modifications related to chromosome silencing and elimination during male meiosis in Bengalese finch // Chromosoma. 2014. V. 123. V. 3. P. 293–302.
Itoh Y. & Arnold A. P. Chromosomal polymorphism and comparative painting analysis in the zebra finch // Chromosom Res. 2005. V. 13. N. 1. P. 47–56.
Kazazian H. H. Jr. Mobile elements: drivers of genome evolution // Science. 2004. V. 303. N. 5664. P. 1626–1632.
Kinsella C. M., Ruiz-Ruano F. J., Dion-Côté A-M. et al. Programmed DNA elimination of germline development genes in songbirds // bioRxiv. 2018. doi:10.1101/444364.
Kumar S., Stecher G., Suleski M. & Hedges S. B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times // Mol Biol Evol. 2017. V. 34. N. 7. P. 1812–1819.
Pigozzi M. I. & Solari A. J. Germ cell restriction and regular transmission of an accessory chromosome that mimics a sex body in the zebra finch, Taeniopygia guttata // Chromosome Res. 1998. V. 6. N. 2. P. 105–113.
Pigozzi M. I. & Solari A. J. The germ-line-restricted chromosome in the zebra finch: recombination in females and elimination in males // Chromosoma. 2005. V. 114. N. 6. P. 403–409.
Torgasheva A. A., Malinovskaya L. P., Zadesenets K. S. et al. Germline-restricted chromosome (GRC) is widespread among songbirds // Proceedings of the National Academy of Sciences. 2019. V. 116. N. 24. P. 11845–11850.
В публикации использованы фото зебровых амадин М. Кулешина и остальных птиц – Н. Андреенковой, О. Андреенкова и Е. Шнайдер



















